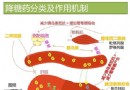
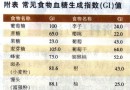
细胞凋亡[21],其机制可能为通过p53功能的丧失,抑或干扰阻断p53与TBP亚基TFIIH及p62亚基TFIID的联系[22].Ghebranious et al[23]通过p53 246ser突变的转基因鼠与表达HBV的转基因鼠进行繁殖,并应用黄曲霉毒素B1作用于后代,结果显示13月龄的p53异常伴有HBV转基因小鼠的HCC发生率高达62%~100%,远较单纯p53突变或HBV转基因小鼠的癌变率高,提示p53突变,HBV感染与黄曲霉毒素在HCC发病中有重要作用.然而,Oguey et al[24]对鼠肝细胞系AML12进行X基因与p53 246ser突变基因的共转染研究,结果并未发现明显的恶性转化,这或许与AML12细胞系对p53突变及pX的作用不甚敏感有关. 4 X基因与pX的表达 HBV DNA整合入宿主细胞基因组是HCC遗传学研究的重大发现之一.目前已知在HBV慢性感染的病程中,HBV DNA以低频率随机地整合入肝细胞基因组,在进行整合的同时常伴随宿主DNA的缺失、重排与转位,最终引起染色体畸变或结构异常[25].在宿主肝细胞基因组,HBV DNA的整合部位不是单一的,它们随机地分布于多条染色体上.然而,统计分析表明这种整合多发生在HBV基因组的X基因区域,因此X基因及其产物pX的表达研究引人注目.在肝癌组织中,X基因与pX表达的阳性率约为50%~90%[26,27].pX主要位于胞质内,部分位于胞核内,且癌周组织的阳性率与癌内相近,但pX在肝癌的表达远较乙肝后肝硬变及慢性乙肝为高,在分离培养的人肝癌细胞仍保持表达pX的特性.此外,约有40%~60%表达X基因或pX的肝癌,其HBsAg的表达为阴性[28] ,提示部分血清HBsAg阴性患者其肝癌组织中仍有X基因的整合.pX在肝癌的表达亦常伴有p53的突变[29].由于X基因的整合与pX的表达既可见于癌细胞,也可见于部分尚无明显癌变的肝细胞或无症状HBV携带者的肝细胞中,因此整合本身仅可能为致癌的起始因素.通过整合可使DNA结构异常,在有辅助致癌因素的作用下可启动癌基因的调节顺序,或通过X基因的反式激活作用,使癌基因转录活性增强而最终导致肝细胞恶性转化.有鉴于此,有学者认为pX的表达可视为HCC发生的先兆.有关pX的血清学检测研究显示无论在HCC抑或慢性乙肝患者的血清中,pX的检出率均较其他乙肝标志物为低,仅为10%~20%[30].这可能因pX是一个弱抗原,或者pX仅在细胞融解时才被释放,因此肝癌细胞内pX表达率与血清pX检出率不呈平行关系.HCC的流行病学调查与实验研究均表明HBV在HCC的发病中具有重要作用.然而,可长达30a的潜伏期也显示HCC的发生是一个多步骤过程.虽然目前已知HBV X基因和pX与肝癌的关系密切,但迄今为止还有许多问题需要进一步研究,这包括X基因的反式激活机制,与宿主肝细胞DNA的整合以及与p53基因的关系等.有理由相信随着X基因的深入研究,将会大大促进肝癌的防治水平.
上一页 [1] [2]