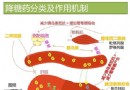
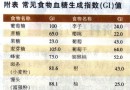
肝肿瘤;乙型肝炎病毒;X基因中国图书馆分类号R735.7 Subject headings liver neoplasms; hepatitis B virus; X gene目前公认肝癌(HCC)的发病是多因素协同作用的结果,其中乙肝病毒(HBV)感染是HCC的主要病因之一.流行病学调查表明HBV感染者HCC的发病率较对照人群高出200倍以上.虽然HBV的确切致癌机制尚未完全阐明,但随着分子病毒学的研究进展,人们对HBV X基因及其产物X蛋白(pX)与HCC之间的关系已有了多方面及多层次的了解,积累了大量研究资料. HBV基因组是一不全双链的环状DNA分子,其长链含有4个开放读码框架(ORF),分别为编码前S1蛋白,前S2蛋白及S蛋白的S基因,编码前C蛋白与C蛋白的C基因,编码DNA聚合酶蛋白的P基因,以及编码pX的X基因.X基因是HBV DNA中最小的一个ORF,编码基因区位于1374~1818核苷酸之间,其编码的pX(或称为乙肝病毒X抗原,HBxAg)由154个氨基酸组成.Kwee et al[1]研究表明X基因至少编码Mr 17000,8000和6600三种蛋白,基因分析显示靠近5'的AUG起始密码开始的序列表达完整长度的Mr 17000蛋白,即pX.Mr8000和Mr 6600两种蛋白是由框架内起始密码子编码的两种缺失氨基端部分序列的pX.三种蛋白在调控中的作用有所不同,只有Mr17000 pX能激活NF-kB依赖性启动子序列,而对SV40的增强子/启动子序列的转录激活,Mr17000和Mr6600蛋白均为必需.可见X基因编码的三种蛋白在功能上互相联系,具有多种转录调节活性. 1X基因的转染与转基因研究X基因的转染与转基因研究使人们对X基因的致癌作用有了突破性认识.1989年Shirakata et al首先发现通过X基因片段的表达载体转染小鼠成纤维细胞系NIH3T3诱发其体外转化,经裸鼠移植可在裸鼠体内形成肿瘤.其后Luber et al[2]应用X基因转染小鼠肝细胞系FMH202,发现所有转染成功的细胞克隆在软琼脂培养中出现恶性生长,形成的细胞集落移植到裸鼠后形成瘤灶.作者认为X基因的表达及肝细胞的基因突变是导致HCC发生的主要原因.X基因转基因研究中最引人关注的是Koike et al[3]将X基因导入CD-1小鼠系的胚胎细胞中,诱发84%的雄鼠肝脏肿瘤并伴有pX的高度表达.实验小鼠在2月龄时围绕中央静脉周围的肝细胞出现胞质空泡样变,伴有pX的高度表达,13月龄时小鼠出现癌灶.经5-溴脱氧尿核苷标记分析,病灶中肝细胞DNA的合成较对照明显增加,且DNA含量分析显示在癌灶出现前转基因鼠的肝脏存在数个非整倍体峰.转基因鼠的研究为X基因在HCC发病中的作用提供了直接的证据. 然而,并非所有的转基因研究均能诱发恶性转化.Reifenberg et al[4]将X基因导入C57B/6J小鼠的胚胎中,经长达2a的观察后处死小鼠,组织学检查显示转基因小鼠的肝脏虽有X基因表达,但病理改变轻微,并无HCC的发生.Balsano et al[5]的研究同样未能在转基因鼠中诱发HCC.对此,Koike[6] 认为能否在转基因鼠中诱发HCC的关键是pX的持续高水平表达,而仅低水平的pX表达不足以导致HCC的发生. 2 X基因的反式激活作用1987年Twu et al发现与PUC 9重组的X基因在猴肾细胞中可激活人β-干扰素基因的调控序列,使受此调控序列调控的氯霉素乙酰转移酶转录增加,从而揭开了X基因反式激活作用研究的序幕.现已明确X基因的反式激活作用具有明显广谱性,pX作为一非特异性反式激活因子,对多种细胞与病毒的多种靶基因均具有反式激活作用[7,8].目前已知pX对多种癌基因具有反式激活作用,其中包括c-myc,ras,c-fos及c-jun等.此外,pX亦能激活胰岛素样生长因子受体Ⅰ,Ⅱ[16,17]及多药耐药基因.因此,人们有理由相信X基因通过其反式激活作用改变宿主基因的表达,导致HCC的发生.pX的反式激活机制非常复杂,主要是以细胞因子作为桥梁,通过复杂的信号传导间接发挥其广谱转录激活功能.与pX反式激活有关的细胞因子有AP-1,AP-2,NF-kB,EF-C及ATF/CREB等.目前已知pX可引起内源性蛋白激酶C(PKC)活性物质DAG增加,激活PKC后引起AP-1,AP-2及NF-kB等细胞因子活化,从而对靶序列发挥激活作用[9].另一方面,pX亦可通过激活Ras-Raf-MAP激酶链锁反应,引起AP-1及NF-kB活化,继而发挥转录激活作用[10].此外,亦有研究显示pX可直接作用于核内组分引发转录激活[11],或者与细胞内蛋白酶、蛋白酶复合物组分[12]抑或DNA修复蛋白[13]发生作用,从而发挥其反式激活作用.由此可见pX的反式激活存在多种途径与方式.Arii et al[14]对pX的一级结构进行分析,发现pX的反式激活功能需要三个关键部位;第46~52氨基酸(特别是46脯氨酸、49组氨酸与52组氨酸)、第61~69氨基酸(特别是61半胱氨酸、67甘氨酸、68脯氨酸与69半胱氨酸),以及第132~139氨基酸(特别是132苯丙氨酸、137半胱氨酸与139组氨酸),这三段序列在不同的嗜肝病毒中高度保守.第46~52氨基酸是形成组氨酸/天冬氨酸特征性功能结构所必需,而第61~69氨基酸及第132~139氨基酸则与丝氨酸蛋白酶抑制有关的Kunitz区域类似,通过此相似结构pX可直接抑制丝氨酸蛋白酶,从而阻断丝氨酸蛋白酶的作用[15].因此,这两种功能结构对pX的反式激活功能至关重要,而第5~27氨基酸及第144~154氨基酸的缺失并不影响pX的反式激活功能.虽然目前对pX反式激活的途径及功能结构有所了解,但由于其作用机制的复杂性,人们尚缺乏统一的认识. 3 X基因与p53基因p53基因是一种肿瘤抑制基因,野生型p53具有维持细胞正常生长,抑制细胞恶性转化的作用.当细胞因物理或化学因素作用发生DNA损伤时,可通过p53启动细胞凋亡过程,使有癌变倾向的细胞不再存活.p53基因的丢失或突变失活不但使其失去野生型p53的作用,而且可能具有癌基因的功能.1991年Hsu et al与Bressac et al分别报道了中国启东和南非地区的HCC,发现在p53的249编码区第三密码子有一特征性点突变G→T颠换(249ser),使原先的精氨酸被丝氨酸所代替,其突变率高达50%,这一结果曾被视为肝癌与p53关系研究中的重大突破.但随后的研究表明此特征性G→T颠换主要由黄曲霉毒素B1引发,而与是否存在HBV感染关系甚微[16].此外,与HCC有关的p53突变尚有249met及220cys[17],但似与HBV感染的联系并不密切.在HBV与p53基因关系研究中引人注目的结果是发现了pX有与P53蛋白结合的作用[18].由于pX与P53蛋白的结合可在蛋白水平使靶细胞中抑癌基因及其产物的功能发生改变,破坏了p53基因的负调节作用,从而导致细胞的恶性转化[19,20].现已证实pX能有效阻断p53介导的
[1] [2] 下一页